会员投稿
会员投稿 手机版
手机版")
")
")
")
")
")
")
非洲猪瘟疫苗的最新研究进展(综述)(上篇)
张红亮,赵赛赛,等
摘要:非洲猪瘟是由非洲猪瘟病毒(African swine fever,ASFV)引起的猪的一种致命传染病。目前,该疾病被列为必须向世界动物卫生组织(World Organization for Animal Health,WOAH)报告的法定通知疾病。自猪瘟暴发以来,全球养猪业遭受的经济损失难以弥补。在当前的疫情期间,控制和根除ASF至关重要。接种疫苗是预防和控制ASF流行的最佳策略,但由于ASF灭活疫苗产生的免疫保护作用较差,并且没有足够的细胞系进行有效的体外ASFV复制,仍有待开发具有高免疫保护潜力的ASF疫苗。了解疾病演变过程、病毒传播方式和疫苗设计突破点,将有助于ASF疫苗的开发。本文旨在重点介绍近年来在ASF的流行和传播、病毒突变以及疫苗开发方面的最新进展和突破,并重点关注未来的方向和趋势。
关键词:非洲猪瘟病毒;流行与传播;亚单位疫苗;DNA疫苗;进展,综述
本文摘译自《Frontiers in Microbiology》,27 April,2023
1 介绍
非洲猪瘟是由非洲猪瘟病毒(African swine fever,ASFV)引起的猪的一种高度传染性疾病,其唯一的天然宿主是猪(Wang等,2019)。它也是唯一感染家猪、野猪和钝缘蜱的大型双链DNA病毒(Dixon等,2019)。该病毒具有直径为200 nm的二十面体对称性和同心圆(Rowlands等,2009)。此外,ASFV基因组包含数量多变的开放阅读框(open reading frames,ORF),范围从160到175个,其中大约125个保守ORF编码50多种功能蛋白(桑切斯-比斯坎诺等,2015a;西莫斯等,2019)。ASF的临床症状分为急性型、亚急性型和慢性型。急性ASF发病迅速,持续时间短,其特征是高烧、食欲不振、发绀、严重内出血和接近100%的死亡率(Pietschmann等,2015;Gaudreault等,2020),而毒力较弱的毒株引起的临床症状较轻。ASF起源于20世纪20年代的非洲,该疾病于2007年传播到格鲁吉亚的高加索地区。从那里开始,ASFV逐渐传播到邻国(即亚美尼亚、阿塞拜疆、俄罗斯和白俄罗斯),影响了家猪和野猪(Rowlands等,2008;科斯塔德等,2009)。自2018年8月以来,ASFV在中国被检测为基因Ⅱ型高毒株(Ge等,2018;Wen等,2019;Zhao等,2019)。辽宁、内蒙古和黑龙江等东北地区先后发生了40多起疫情,给中国养猪业造成了前所未有的损失,并严重影响了国民生产和饮食结构。随着经济和国际贸易的持续增长,近年来已采取重大措施遏制病毒传播。了解ASFV的全球形势、流行情况和传播途径对于预防疾病暴发至关重要。
1957年,ASFV基因Ⅰ型被称为ESAC-WA因型,此后传播到欧洲、南美和加勒比海地区;到2007年,所有国家(除意大利和非洲外)都宣布根除了ASF。自2007年ASFV基因Ⅱ型暴发以来,世界再次暴发ASF,包括俄罗斯(2007)、乌克兰(2012)、波兰(2014)、比利时(2018)、中国(2018)、越南(2019)、印度(2020)和许多其他国家。除了传播范围广之外,传播途径也非常多样。ASF是一种通过直接或间接接触受感染的猪和粪便传播的病毒性疾病。蜱等生物媒介也加速了病毒的传播。车辆、工具和人类活动等机械因素,以及生猪加工产品也是致病因素。生物防治是目前预防ASF最直接有效的方法,优于接种疫苗。
ASF灭活疫苗已被证明对预防ASF无效。令人惊讶的是,其中一种减毒活疫苗ASFV-G-δI177L是世界上第一种商业化的ASF疫苗(Tran等,2022a)。然而,减毒活疫苗的特点是研发成本高、周期长、不稳定,这限制了其快速发展。研究人员目前专注于基因工程疫苗,如亚单位疫苗、载体疫苗和DNA疫苗,而大多数实验仅限于免疫原性,没有进行攻毒实验。因此,开发安全有效的ASF疫苗是对抗该病毒的首要任务之一。本综述重点阐述了ASF疫苗的最近进展和优缺点,并为有未来的研究提供了新的方向。

如有需要,请点击上述图片
2 ASF的全球情景和流行率
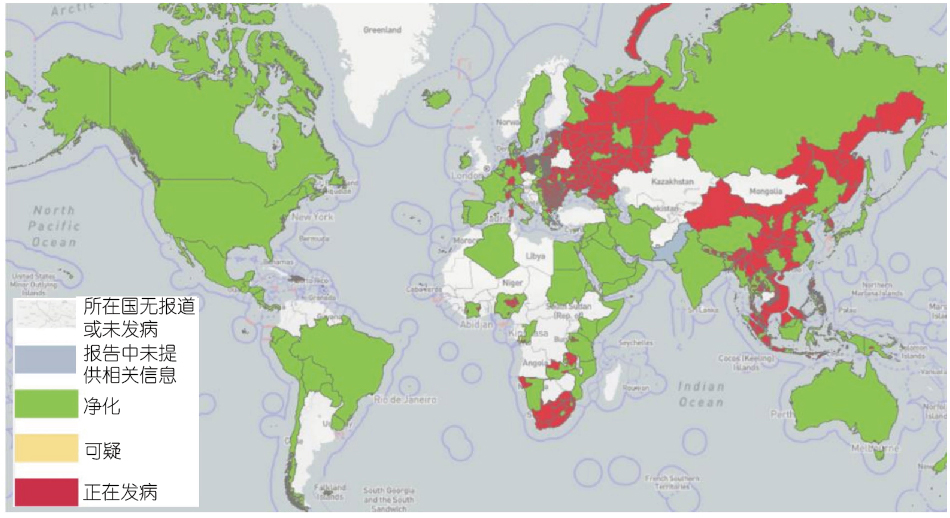
非洲猪瘟原产于非洲,它通过一种古老的森林循环来维持病毒,这种循环涉及疾病的天然宿主和传播媒介,以及有或没有天然病媒参与的家畜循环(穆伦巴-姆富穆等,2019),自2005年以来,32个国家报告暴发了ASF。在撒丁岛,流行着ASFV基因Ⅰ型,它仍然是呈地方性流行(Franzoni等,2020),而ASFV基因Ⅱ型引起的疫情于2007年在格鲁吉亚暴发,并从那里传播到邻国,包括亚美尼亚、阿塞拜疆、俄罗斯和白俄罗斯;在那里,ASFV用其高毒力的分离株感染家猪和野猪(Karger等,2019)。2014年至2018年,欧洲大陆欧盟共有9个国家,如爱沙尼亚、立陶宛、拉脱维亚、波兰、捷克共和国、保加利亚、比利时、罗马尼亚和匈牙利,受到ASF的严重影响,尽管努力控制ASF,但ASFV仍持续传播。野猪感染ASFV后会出现高病死率,是典型的急性和亚急性感染形式,特别是在新感染地区(Martínez-Avilés等,2020)。从2020年到2022年的短短2年时间里,共有16个欧盟国家报告了这种疾病。值得注意的是,比利时(2020年3月)和捷克共和国(2018年4月)成功根除了ASF巴诺斯等,2022)。2018年8月,ASF在中国首次得到证明(Liu等,2021),并在几个月内迅速蔓延到31个省份,导致总共178起ASF疫情。在2019/2020年,该疾病蔓延到大洋洲,东帝汶报告了非洲猪瘟,导致小型猪场受到受到感染后猪出现了很高的的死亡率(Berends等,2021;Phillips等,2021)和巴布亚新几内亚(Mighell和Ward,2021)。2020年初,印度(阿鲁纳恰尔邦和阿萨姆邦)首次出现高死亡率的ASF,这与亚洲和欧洲报告的2007年后ASFV p72基因Ⅱ型病毒相似,表明该地区暴发的猪瘟具有跨境感染趋势(Rajukumar等,2021)。2021年7月,ASF在消失近40年后再次出现在美洲,先是在多米尼加共和国,然后在海地,威胁到猪的健康、牲畜市场和生产者的生计(Jean-Pierre等,2022)。同样,在2022年1月,意大利大陆也在缺席约40年后通知出现了ASFV基因Ⅱ型(Beato等,2022;Iscaro等,2022)。与此同时,两个新的国家受到了影响:北马其顿的两个国家之一,是由以小规模为主的农场造成的,这些农场人员流动率高,野猪出现的频率最高(O'Hara等,2021),另一起发生在泰国,由受感染的活猪和死猪、猪肉制品以及野猪精液、受污染的饲料和污染物(Thanapongtharm等,2022)。尼泊尔报告的首例非洲猪瘟发生在2022年3月,尽管尼泊尔政府已于2019年1月28日禁止从感染ASFV病毒的国家进口猪和猪肉产品(Acharya和Wilson,2020;图1)。
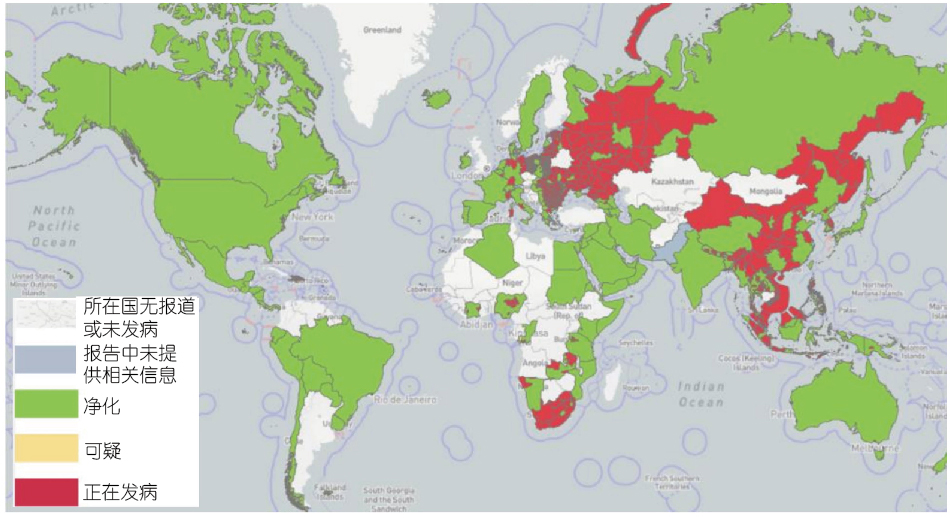
图1 自2020年以来,截至2022年9月29日,全球已有45个国家报告了ASF疫情(来源:世界动物卫生组织、世界卫生组织,2022年10月线访问)。

如所示表1,自2020年1月以来,五个不同地区报告了暴发ASF,涉及45个国家,影响了超过1 129 000头猪和36 000头野猪,造成超过1 931 000头猪死亡(数据来自即时通知和后续报告)。ASF在全球的暴发导致全球生猪产能急剧下降,养猪业遭受重创。由于ASFV感染的隐匿性和复杂性,目前疫情仍不明朗。预计在3~5年内很难恢复到以前的水平。

3 ASFV的传播途径
ASFV具有广泛的传播途径,包括直接和间接传播,导致预防和控制ASF变得复杂化(图2)。野猪和蜱是ASFV的天然宿主,在森林循环的传播中发挥着关键作用(Sánchez-Vizcaíno等,2015a;mietanka等,2016;Galindo和Alonso,2017)。无处不在的野猪群对在ASF发生的地区控制和根除ASF构成了重大障碍。发病和死亡的野猪是ASFV欧洲毒株的主要来源,其对不同的ASFV基因型易感,特别是ASFV基因Ⅱ型分离株(Blome等,2012),是直接或间接传播疾病的重要媒介(Probst等,2017)。先前的研究表明,ASFV可以通过野猪和家猪传播,这加剧了病毒在家猪和野猪以及同品种种群之间的传播(Costard等,2013)。在家养猪循环中,家猪是病毒的唯一宿主和携带者,病毒一直保持被感染,包括猪的粪便和分泌物(Guinat等,2016)。事实上,其他动物,如苍蝇、水蛭和鸟类,包括秃鹫,也可能通过远距离机械传播,成为病毒携带者(Brown和Bevins,2018;Olesen等,2018;Karalyan等,2019;Bonnet等,2020)。

此外,人类活动促使ASFV流入猪场,加剧了病毒传播的程度。ASFV传播的一个重要途径是通过非法运输受到感染的猪肉产品和受到污染的物品,包括饲料、设备、车辆和衣物(Costard等,2013)。流行病学的其他关键因素,包括运输受到污染的猪肉产品和饲喂泔脚,已在高加索、俄罗斯联邦和中国(Gogin等,2013)。ASFV的固有特性对多种环境条件(如温度、pH等)具有高度抵抗力,导致病毒确实存在于环境中,并持续传播到遥远的地区(Davies等,2017;Schulz等,2017;Mazur-Panasiuk等,2019;Petrini等,2019;Zani等,2020)。有趣的是,多项研究证明,在特定条件下,ASFV可以在跨大西洋航线上的动物饲料中存活(Dee等,2019;Stoian等,2019)。此外,尽管受到病毒污染的冷冻猪肉和加工猪肉制品不会导致人类感染,但由于病毒的长期存活,它们可能是病毒在长途运输后传播到新地区的潜在因素(Schulz等,2017)。
4 ASFV的遗传变异
随着ASFV分型技术的持续发展,对ASFV基因组遗传进化的研究进入了一个新阶段;到目前为止,ASFV已被分为24种基因型(Achenbach等,2017)或8个血清群(Malogolovkin等,2015)。对ASFV基因分型的深入研究有助于在分子水平上追踪引起疫情的病毒来源,了解潜在的传播途径和可能的传播方式,并筛选潜在的候选疫苗株。

图2 ASFV病毒向猪的潜在传播途径。
ASFV基因组是线性双链DNA,包括左侧可变区(variable regions,LVR)、中央保守区(central conserved region,C区)和右侧可变区(right variable region,RVR。Van Etten,2009)。不同毒株的基因组序列可能在位点上有显著差异,例如,LVR内的多基因家族(MGF)、中央可变区(CVR)和C区内的EP402R基因(表达CD2v蛋白)。不同的地区为ASFV的进化分析提供了有利的条件,特别是遗传进化。
4.1 ASFV基因I型毒株
ASFV基因Ⅰ型毒株于1957年在非洲以外的葡萄牙首次发现并引起暴发(Revilla等,2018)。此后,在20世纪50年代至90年代期间,该毒株在葡萄牙、西班牙、古巴、巴西和其他国家流行(Sánchez-Vizcaíno等,2015b;Galindo和Alonso,2017)。作为亚洲地区报告的首例病例,2021年首次在中国临床样本中检测到ASFV基因Ⅰ型毒株(Sun等,2021)。目前,ASFV基因Ⅰ型毒株仍在一些西非国家占主导地位,而北非国家没有报告暴发ASF疫情(Mulumba-Mfumu等,2019)。在中非,ASFV基因Ⅰ型和Ⅱ型毒株同时占主导地位。表2列出了ASFV的具体毒株信息。
葡萄牙等国家的科研人员对ASFV的高毒力毒株lisbo a60(L60株)和低毒力毒株NH/P68株(NHV株)的基因组进行了测序,将差异部分与ASFV的其他已知毒力的毒株进行了比较。分析表明,ASFVL60株和NHV株与来自欧洲和西非的p72基因Ⅰ型ASFV毒株最近相关,支持了欧洲毒株起源于西非的假设。由于L60和NHV之间的长间隔和地理距离,暗示这是由于葡萄牙分离物在组织培养中的广泛传播(葡萄牙等,2015)。该团队比较了ASFV高毒力lisbo a60株(L60株)和低毒力NH/P68株(NHV株)的基因序列,以克服病毒-宿主相互作用研究的限制,揭示了对病毒进化的进一步了解(Portugal等,2020)。
自2007年ASFV基因Ⅱ型毒株在格鲁吉亚出现以来,世界已陷入一场对抗ASFV基因Ⅱ型株流行病的战争。ASFV基因Ⅰ型毒株与目前流行的基因Ⅱ型毒株有很大不同。从山东和河南猪场的临床猪标本中分离到两株不具有猪红细胞吸附活性的ASFV基因Ⅰ型SD/DY-I/21株和HeN/ZZ-P1/21株。全基因组分析显示,这两种毒株与20世纪在葡萄牙分离到的ASFV基因Ⅰ型低致死性毒株NH/P68株和OURT88/3株的高度相似,与早年在欧洲和非洲分离到的ASFV基因Ⅰ型高致病性毒株L60株和Benin 97株的有显著差异。尽管全基因组序列高度相似,但ASFV基因Ⅰ型SD/ DY-I/21株和HeN/ZZ-P1/21株在某些基因上存在有意义的差异,这表明它们可能属于不同的入侵来源(Sun等,2021)。ASFV基因Ⅰ型和基因Ⅱ型毒株在中国的共存,凸显了当前防控形势变得更加严峻,加速了疫苗的开发。

4.2 ASFV基因Ⅱ型毒株
自从基因克隆、PCR和测序技术迅速发展以来,ASFV基因组研究取得了令人满意的进展。p72基因分型是目前广泛用于ASFV基因分型的金标准。Gonzague等证实,1999年分离的ASFV马达加斯加毒株与1994年分离的ASFV莫桑比克毒株有99.2%的相关性,具有p72基因的高度保守片段(贡扎格等,2001)。2003年,Bastos等首次使用p72蛋白的C端将ASFV分为10种基因型(Bastos等,2003)。2005年,Lubisi等(2005)进一步将ASFV分为16种基因型。2007年,Boshoff等对43株ASFV分离株(1973—1999)进行了研究(Boshoff等,2007)。2016年,Achenbach等(2017)对埃塞俄比亚猪群中分离到的ASFV(2011年至2014)进行了比较分析,并再次确定了一种基因型。2017年,Quembo等(2018)利用从莫桑比克戈龙戈萨国家森林公园采集的软蜱样本中分离出的19种病毒株分析了ASFV病毒。进化树特异性分析显示,5个毒株属于一个新的进化分支。因此,总共24种ASFV基因型被完全鉴定。
为了更好地了解相似毒株的进化趋势,Gallardo等研究发现,ASFV的整个基因组在I73R和I329L基因的基因间区(intergenic region,IGR)有不同的串联重复序列(tandem repeat sequences,TRS)组合。例如,从波兰和立陶宛分离到的ASFV分离株的IGR与从白俄罗斯和乌克兰分离到的ASFV分离株的IGR相同,但与ASFV俄罗斯分离株的IGR不同,这表明在波兰和立陶宛流行的ASFV可能起源于白俄罗斯(Gallardo等,2014)。后来发现,自2012年以来,ASFV的俄罗斯分离株在IGR位点上发生了变化,推断在欧盟流行的ASFV可能来源于俄罗斯。2019年,ASFV的中国/广西/2019株被命名为IGR-III(Ge等,2019),该毒株在IGR位点上有两个串联重复序列,主要在中国流行。随后,该毒株也相继出现在韩国(Kim等,2021)和越南(Nguyen等,2022)。波兰也发现了一种具有三个串联重复序列的病毒株,即IGR-Ⅳ。
2020年,德国ASFV大流行毒株3′端的DNA聚合酶PolX基因O174L可区分为五个不同的品系,至少有10种不同的突变,导致基因的致病作用发生了改变(Forth等,2023)。同年,印度尼西亚兽医科学研究中心收集了西爪哇省茂物地区养猪场的样本。ASF阳性率为16/19,基因组分析表明这种ASFV基因型与在越南、中国和俄罗斯家猪流行的基因型IIASFV分离株的相同(Dharmayanti等,2021)。Senthilkumar等首次揭示了印度的ASFV分离株的基因组分析结果,该分离株与亚洲的流行毒株关系密切。有助于识别亚洲流行株的一个重要特征是I73R和I329L基因之间的错配(Senthilkumar等,2022)。2021年,对在沙巴流行的ASFV分离株的基因进行分型显示,该毒株与中国、越南和印度尼西亚的流行毒株相当(Khoo等,2021)。
因此,未来应重视在全基因组水平上监测ASFV基因组,这有利于掌握ASFV的流行病学模式,研究ASF演变,筛选理想的候选疫苗株。
未完,待续